***We are looking for camera-trap carnivore records from the dry and moist forests of Western Ecuador and Peru! Please see below and contact us for more information! ***
Motivation
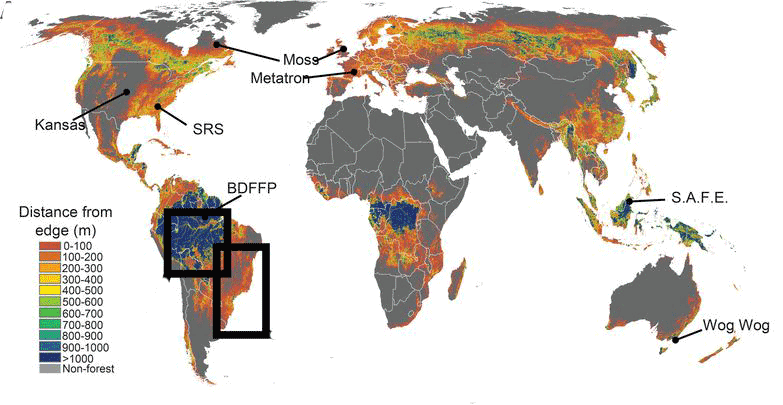
Map of fragmented landscapes showing distance from the forest edge and, indirectly, patch size. Large patches are shown in blue and small patches in red and orange. (Haddad et al. 2015)
Currently, more than a third of the global forest cover has been lost because of deforestation and land cover change 1, leaving only about 7.5% of global protected areas connected 2. This pervasive habitat loss has contributed to the isolation of protected areas and forest patches which exacerbates the extinction risk of endangered populations 3,4.

(c) Jonathan Truong
The groups most affected by patch isolation are apex predators and other large vertebrates 3,5,6. Species within the order Carnivora increase their extinction risk after habitat loss and patch isolation because of their large spatial needs and low population densities 6. Additionally, cascading effects after carnivore extinctions and range contractions have several negative effects on community structure and food-web dynamics, such as herbivore hyper-abundance and meso-predator release 7,8. Thus, it is imperative to identify the factors that affect carnivore persistence in isolated populations and protected areas 6,9,10.
To understand carnivore connectivity, and identify corridors to improve connectivity, descriptions of both landscape structure and species-specific habitat suitability are needed 11. Habitat suitability information in connectivity studies is usually converted to a resistance surface as a measure of the estimated cost for an animal to transverse that area 12. A rigorous but data-intensive approach to corridor planning is to develop empirical habitat suitability models from field surveys. Methods to obtain habitat suitability information for large carnivores include camera trapping surveys 13, radio- or GPS-collar telemetry data 14, and other indirect evidence (i.e. scat sampling) 12,15. The resulting information on carnivore-habitat relationships can be used in corridor designs based on least-cost path models or circuit theory 16,17.
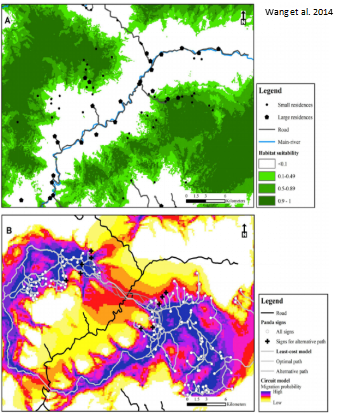
A habitat suitability map showing suitable areas in dark green. Below: Circuit model showing high and low suitable areas for corridors. (Wang et al. 2014)
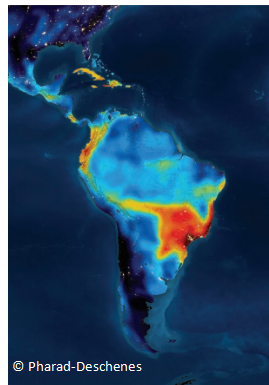
A heat map showing the number of species at risk. More than 100 species of vertebrates at risk are shown in dark red while zero species at risk is shown in dark blue.
The Project
The two largest areas of deforestation, high human pressure, and biodiversity loss in South America are located in the Atlantic Forest in eastern Brazil and northern Argentina, and in the Pacific Forests of northern South America. Because the latter holds critically endangered ecosystems, such as the Tumbesian dry forests, it had been proposed as the Tumbes-Choco-Magdalena hotspot (TCMH) for conservation priority. Thus, we will work in the southern forests of Ecuador and Peru, within the TCMH. Currently, our main objectives are to: 1) identify the factors affecting carnivore persistence in fragmented landscapes and 2) identify species-specific responses to fragmented landscapes.
From the Field
Thanks to NGO’s Spectacled Bear Conservation Society and BioS for sharing their data and collaborating with us! We are also joining efforts with the University of Guayaquil and in the next few weeks we will start to set cameras in Isla Santay near Guayaquil City in Ecuador!! We are currently gathering existing field data sets to include in our analysis. So if you have carnivore records and would like to collaborate, please contact us at cindymel[at]mail.ubc.ca and cole.burton[at]ubc.ca!
References:
- Hansen, M. C. et al. High-resolution global maps of 21st-century forest cover change. Science 342, 850–853 (2013).
- Saura, S. et al. Protected area connectivity: Shortfalls in global targets and country-level priorities. Biological Conservation 219, 53–67 (2018).
- Laurance, W. F. et al. Averting biodiversity collapse in tropical forest protected areas. Nature 489, 290–294 (2012).
- DeFries, R., Hansen, A., Newton, A. C. & Hansen, M. C. Increasing isolation of protected areas in tropical forests over the past twenty years. Ecological Applications 15, 19–26 (2005).
- Crooks, K. R. Relative sensitivities of mammalian carnivores to habitat fragmentation. Conservation Biology 16, 488–502 (2002).
- Crooks, K. R., Burdett, C. L., Theobald, D. M., Rondinini, C. & Boitani, L. Global patterns of fragmentation and connectivity of mammalian carnivore habitat. Philosophical transactions of the Royal Society of London. Series B, Biological sciences 366, 2642–51 (2011).
- Ripple, W. J. et al. Status and ecological effects of the world’s largest carnivores. Science 343, 1241484 (2014).
- Prugh, L. R. et al. The Rise of the Mesopredator. BioScience 59, 779–791 (2009).
- Regolin, A. L. et al. Forest cover influences occurrence of mammalian carnivores within Brazilian Atlantic Forest. Journal of Mammalogy 98, 1721–1731 (2017).
- Michalski, F. & Peres, C. a. Anthropogenic determinants of primate and carnivore local extinctions in a fragmented forest landscape of southern Amazonia. Biological Conservation 124, 383–396 (2005).
- Keeley, A. T. H., Beier, P. & Gagnon, J. W. Estimating landscape resistance from habitat suitability: effects of data source and nonlinearities. Landscape Ecology 31, 2151–2162 (2016).
- DeMatteo, K. E. et al. Using niche-modelling and species-specific cost analyses to determine a multispecies corridor in a fragmented landscape. PLOS ONE 12, e0183648 (2017).
- Brodie, J. F. et al. Evaluating multispecies landscape connectivity in a threatened tropical mammal community. Conservation Biology 29, 122–132 (2015).
- LaPoint, S., Gallery, P., Wikelski, M. & Kays, R. Animal behavior, cost-based corridor models, and real corridors. Landscape Ecology 28, 1615–1630 (2013).
- Angelieri, C. C. S. et al. Using species distribution models to predict potential landscape restoration effects on puma conservation. PLoS ONE 11, 1–18 (2016).
- Sawyer, S. C., Epps, C. W. & Brashares, J. S. Placing linkages among fragmented habitats: do least-cost models reflect how animals use landscapes? Journal of Applied Ecology 48, 668–678 (2011).
- McRae, B. H., Dickson, B. G., Keitt, T. H. & Shah, V. B. Using circuit theory to model connectivity in ecology, evolution, and conservation. Ecology 89, 2712–2724 (2008).